

![]()
Cerebros femeninos y masculinos: del talento a la combatividad
La noción más concurrida, hoy en día, sobre los hiatos de género es que las diferencias cognitivas entre hombres y mujeres son nimias y del todo prescindibles. El acceso femenino a todas las esferas de la actividad profesional ha sido tan abarcador y formidable que cuesta sustraerse a esa certidumbre. Se da por descontado un horizonte de igualdad taxativa para ambos géneros en cualquier aptitud y actividad, y se obvia el detalle “menor” de que algunos sesgos persistentes en las preferencias y aficiones puede que deriven de propensiones enraizadas en la biología de la diferenciación sexual. Aquella prescripción igualitaria gravita, sin embargo, sobre un error edificado a partir del descarte de un vasto arsenal de hallazgos sobre el particular. En realidad, las diferencias anatómicas y fisiológicas entre los varones y las féminas humanas son prominentes tal y como suelen serlo en el resto de mamíferos. Y también lo son las vivencias afectivo/emotivas y los perfiles de rendimiento motor, sensitivo y cognitivo. Los distingos entre sexos afectan a prácticamente todos los sistemas y engranajes del organismo y dependen de mecanismos intrincadísimos. Eso también vale para el cerebro, que es la glándula donde suelen sustanciarse los habituales y no siempre benignos conflictos de género.
Todo ello no impide constatar que las conquistas femeninas en cualquier capacitación, así como en las gradaciones jerárquicas en empresas e instituciones, han sido apabullantes a lo largo de las últimas décadas y que los cambios provocados por el poder femenino han resultado no menos impactantes. Persisten, no obstante, algunos reductos profesionales y de esparcimiento menos propicios para el avance de las mujeres. En las disciplinas científicas, por ejemplo, el progreso ha sido meteórico en términos globales: así, en USA, mientras que en 1970 menos del 5% de los doctorados en ciencias y medicina se otorgaban a mujeres, en 2006 ellas se llevaban ya el 51% de los doctorados en biología, el 76% en veterinaria, el 67% en psicología y el 50% en medicina (14,15). Vuelcos radicales aunque esas proporciones se quedan cortas en Europa donde se ha virado hacia un monopolio femenino, en esas áreas, con expulsión casi total de los varones (o deserción apresurada). Existe, no obstante, una excepción: en las disciplinas con un fuerte peso ingenieril o matemático el empuje femenino ha sido menor. En 2006 menos de una tercera parte de los doctorados USA fueron para ellas: 29,6% en matemáticas, 21,3% en ciencias de la computación, 29% en físicas y 20,2% en ingeniería. Según datos extraídos de las primeras 100 Universidades USA, la conquista femenina de los escalafones del profesorado permanente se mueve, en esas disciplinas, entre el 8,8% en ingeniería mecánica y el 15,8% en astronomía (40), y por lo que respecta a los catedráticas apenas alcanzaba la cota del 10%: un 9,7% en química, 7,1% en matemáticas, 10,3% en ciencias de la computación, 6,1% en física, 7,3% en ingeniería química, el 7,1% en ingeniería eléctrica , 7,3% en ingeniería de caminos, 4,4% en ingeniería mecánica y un 8,7% en económicas. El origen de esa discordancia se ha debatido con apasionamiento, con episodios tan conocidos como el cese, en 2006, como Presidente de la Universidad de Harvard, de Larry Summers, un economista de postín y director de la Asesoría Económica en las administraciones Clinton y Obama, por emitir opiniones “incorrectas” sobre el tema.
El viejo dilema de la génesis “natural” (sistema nervioso y endocrino distintivos) versus los efectos de “entornos educativos” separadores (crianzas sesgadas en función de los roles esperables para niñas y niños), se ha enquistado en el meollo de los recurrentes litigios sobre la cuestión de los hiatos y los techos alcanzables por varones y féminas[1]. Por ahí, por esas fronteras irritantes y no clausuradas voy a adentrarme en el asunto, aunque rastrearé otras vías de acceso.
Cuerpos masculinos y femeninos: la impregnación hormonal.
Nadie debate, en cambio, el origen de la espléndida diferenciación morfológica entre hombres y mujeres: los notables contrastes entre el chasis y la estampa varonil en comparación con las ondulaciones femeninas en la plasmación de los ornamentos corporales más peculiares y distintivos. Ahí no hay conflicto, a pesar de la complicación de los estadios intersexuales y del tuneo radical de carrocería que suelen demandar los transexuales que se sienten a íntimamente a disgusto con el molde femenino o varonil donde arribaron, espontáneamente, al madurar. Mediante cirugía correctora y reconstructora, combinada con tratamientos hormonales, pueden viajar desde un patrón corporal a otro, con incomodidades innegables pero con una pulcritud y una resolución escultórica a menudo impecable.
Ahí no hay discusión alguna: el cuerpo es materia modelable y los desajustes se solventan mediante cambios morfológicos (biofísicos) y moleculares (bioquímica hormonal, preferentemente). El papel de las hormonas sexuales en la plasmación del prototipo “atlético” masculino y de las ondulantes siluetas femeninas no admite dudas, vacilaciones o mistificaciones. Se acepta, sin rechistar, que además de la organización distintiva de los genitales, de la distribución ósea, muscular y del tejido adiposo, de la presencia y distribución peculiar del vello, de las cualidades de la piel y hasta de las modalidades tonales distintivas en la emisión de la voz, al hablar o cantar, es el resultado de la acción hormonal trabajando sobre dianas tisulares en etapas singulares del crecimiento. Es oportuno reseñar, por cierto, que el registro diferencial de las voces femeninas y masculinas da lugar a tipologías vocálicas variadas, para ambos géneros en el “bel canto” y en cualquier ámbito musical, sin que ello ocasione conflictos entre los cantantes consagrados, los meritorios o los aficionados. Quiero decir, con ello, que no se conocen reclamaciones ni medidas para propiciar la paridad de géneros entre los barítonos, los bajos, las mezzo o las sopranos, aunque esas y otras especializaciones vocales dependan de sutiles mecanismos de regulación neuromotora distintiva.
Tampoco se conocen aspiraciones serias para alcanzar paridades de género en materia de reacciones sexuales placenteras. Es más, suele ser motivo de celebración gozosa el disfrute de reflejos diferenciados en la respuesta sexual femenina y masculina humana durante la exaltación del juego erótico y la cópula, dando por sentado que las singularidades estentóreas (la erección peneana o la lubrificación vaginal, por poner los ejemplos distintivos más obvios), obedecen a intrincados circuitos neuromotores con su respectiva especialización reguladora, espinal y encefálica (45,59). Una especialización distintiva que también se da para la modulación cerebral del goce, la exaltación o la saciedad amorosa (21). Esa querencia por acentuar, si cabe, las singularidades entre géneros en los ritmos y la hondura del deleite sexual se extiende, asimismo, al abanico de las preferencias en las dianas del interés erótico, y eso requiere una regulación neural mucho más “alta”, por así decir. Alejada de la mediación automática o “instintiva”.
Cerebros dispares: de los moldes a los talentos.
La encrucijada decisiva de los debates sobre hiatos de género suele situarse, no obstante, en el encéfalo más “sofisticado” porque todo el mundo entiende que ahí residen los vectores nucleares del ingenio, el discernimiento y el temple (29,31,34,51). Dado que existen sospechas fundadas y datos abundantes sobre diferencias de talentos y de carácter, entre hombres y mujeres, hay prisa para intentar vincularlas con distingos en la organización y funciones del universo cortical y subcortical. Las primeras aproximaciones a la mensuración precisa en series de cerebros autopsiados y normativos corroboraron, de entrada, que el volumen cerebral femenino es inferior al masculino tanto en términos absolutos como en los relativos en función de la altura o del tamaño corporal. Se trata de una diferencia volumétrica que se mueve entre los 130-200cc, en promedio, a favor de los varones y que no se refleja en agudeza mental ya que en las medidas de ingenio global no hay diferencias consignables entre géneros o, cuando las hay, se decantan ligeramente a favor de las féminas. Es decir, por ponerlo en términos sencillos, que los especímenes humanos de ambos sexos consiguen agudezas cognitivas equivalentes con cerebros de distinto tamaño, con lo cual las diferencias en el volumen total no sirven para explicar los distingos en los talentos y en el temperamento.
Con el inicio del presente milenio proliferaron los mapas comparativos a partir de series de cerebros masculinos y femeninos escaneados mediante Resonancia Magnética. Estudiando 40 cerebros masculinos y 40 femeninos normativos, entre los 18- 45 años de edad, neuroradiólogos de Pennsylvania capitaneados por Rubén Gur (25), confirmaron aquellas diferencias globales usando procedimientos automatizados de segmentación de la sustancia gris, la sustancia blanca y el líquido céfalo-raquídeo. Corroboraron también hallazgos anteriores que indicaban que, en el encéfalo femenino, el porcentaje relativo de sustancia gris es mayor, mientras que en los varones predominan los compartimentos de sustancia blanca y de líquido céfalo-raquídeo. En los hombres, además, consignaron la existencia de asimetrías en esos segmentos encefálicos grises o blancos entre el hemisferio derecho o el izquierdo, unas discrepancias inter-hemisféricas sustanciales que no se daban en las mujeres. Tanto los volúmenes de sustancia gris como los de sustancia blanca se vincularon con los rendimientos cognitivos en una batería de tests neuropsicológicos, confirmando así la relación positiva entre el tamaño de las partes nobles de la glándula craneal y la agudeza y la precisión en el trabajo mental. Esas distinciones en porcentajes volumétricos y en distribución asimétrica del tejido neural puede que medien las diferencias cognitivas entre los dos géneros, y esos investigadores se atrevieron a sugerir que como la sustancia gris se compone de cuerpos neuronales y terminaciones dendríticas, mientas que la sustancia blanca está formada por prolongaciones axonales, quizás el encéfalo femenino suple la disminución de talla e interconectividad a distancia con un aumento de la capacidad computacional en unos circuitos más densos y mejor empaquetados.
Goldstein y col. (23) compararon los volúmenes regionales en 27 cerebros masculinos y 21 femeninos que no diferían en edad, lateralidad manual, inteligencia, educación o nivel socio-económico, teniendo siempre como referencia el volumen total cerebral. Detectaron diferencias entre géneros en bastantes áreas corticales y subcorticales con un patrón femenino denotando un mayor volumen relativo en diversas zonas de la neocorteza, sobretodo en regiones frontales y cinguladas, mientras que los hombres tenían volúmenes superiores en la amígdala, el hipotálamo y la corteza frontal-medial, confirmando así numerosos hallazgos previos obtenidos en estudios anatómicos post-mortem. Esas diferencias se vincularon, asimismo, con las concentraciones regionales distintivas de receptores para los esteroides sexuales, según estimaciones efectuadas en mamíferos, desde roedores hasta macacos, lo cual supone una corroboración indirecta de la influencia de esas hormonas en la organización encefálica diferencial entre ambos sexos.
La Tabla I recoge los valores de comparaciones efectuadas en un escaneo MRI, con procedimientos automatizados de segmentación tisular y zonal, por el equipo de Hanna Damasio, en Iowa, en 23 varones y 23 hembras, normativos, adultos y diestros (1). En todas las estructuras encefálicas, los volúmenes masculinos globales así como los de sustancia gris y blanca fueron superiores a los femeninos, aunque en la proporción G/B (sustancia gris/sustancia blanca) las mujeres dieron, casi siempre, cifras superiores. La distancia entre sexos fue superior para la sustancia blanca que para la sustancia gris y debe remarcarse que buena parte de las variaciones en la proporción entre uno y otro segmento derivaban de la menor presencia de sustancia blanca en las féminas. Ese patrón general se repitió, con ligeras excepciones, en la mayoría de regiones corticales. A pesar de la considerable concordancia con hallazgos previos, ya fuere usando metodologías automatizadas de segmentación o en medidas post-mortem, los autores demandaban cautela al vincular esos dimorfismos sexuales con los patrones distintivos en los rendimientos cognitivos entre ambos géneros.
Tabla I. Volúmenes (cc) de los segmentos cerebrales por regiones con la proporción G/B (sustancia gris/sustancia blanca), las diferencias zonales entre géneros, la magnitud del efecto y la significación estadística de los contrastes. (tomada de 1)

Además de esos contrastes zonales para los segmentos tisulares y del líquido encefálico, también se llevaron a cabo aproximaciones globales estudiando el grosor cortical en muestras normativas con amplios rangos de edad. En un estudio del grupo de neuroradiólogos UCLA que lidera Arthur Toga, ponderaron eso en 176 sujetos con edades comprendidas entre los 7 y los 87 años, además de contrastar diferencias regionales y globales de volumen cerebral (51; Tabla II). El mapeo fino de las diferencias sexuales en grosor cortical reveló que las mujeres tenían una neo-corteza mayor en las regiones parietales inferiores y temporales posteriores. En esas regiones, la cenefa de la sustancia gris era unos 0,45mm. más profunda en las mujeres que en los hombres. Al comparar una sub-muestra adulta formada por 18 hombres y 18 mujeres emparejados por edad se corroboraron esos hallazgos, con independencia de los volúmenes globales y del tamaño corporal. Esa superioridad femenina en sustancia gris temporo-parietal parece sólida e independiente de la edad. En los hombres se constató, sobretodo, una superioridad en los volúmenes de las regiones pre-frontales y occipitales. En trabajos adicionales corroboraron, además, que el cerebro femenino compensa la disminución de talla con un incremento de la complejidad en el plegamiento de la corteza.
Tabla II. Diferencias entre sexos en el volumen cerebral total, en la sustancia gris total, en la sustancia blanca total y en el liquido céfalo-raquídeo total en los hombres (N=90) y mujeres (N=86) de la muestra UCLA de 176 sujetos (tomada de 51)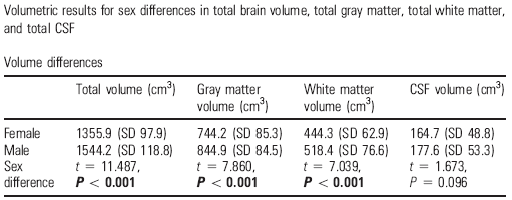
Las comparaciones lo largo del desarrollo y la maduración encefálica han sido culminadas, con esmero, en bastantes estudios dedicados a las criaturas y los adolescentes (35), y también se han contrastado los ritmos de envejecimiento diferencial, según el género. En cuanto a las posibles vinculaciones con medidas de QI, hay que referirse a un estudio pionero en UCLA (26) llevado a cabo con 48 sujetos normativos procedentes de dos muestras, una de adultos jóvenes (edad media = 27+/-6 años) , y otra de personas en la madurez avanzada (edad media = 59 +/- 16 años ). Se analizaron las vinculaciones entre las variaciones estructurales en el cerebro de hombres y mujeres con las notas de QI. La mayor parte de esas relaciones tenían un sentido positivo pero con diferencias notorias entre ambos sexos. En los varones las asociaciones más potentes y amplias entre sustancia gris y QI global se dieron en ambos lóbulos frontales (BA 8, 9) y en el parietal izquierdo (BA 39, 40) es decir, en zonas Wernicke. En las mujeres, en cambio, esos vínculos se restringieron al frontal derecho (BA10) y en particular al área de Broca (BA 44, 45), aunque no fueron tan amplios como en los varones. Por otra parte, en las mujeres hubo mucha más covariación entre sustancia blanca y QI que en los hombres, en varias zonas corticales aunque con predominio frontal, mientras que para la sustancia gris el panorama fue justo el inverso: mayores asociaciones en ellos que en ellas. Los vínculos con las medidas de QI manipulativo o verbal fueron tenues. Por consiguiente, reaparece la conclusión de que mujeres y hombres alcanzan puntajes similares de agudeza cognitiva global con cerebros bastante distintos y con una contribución regional diferenciada, además, por zonas y compartimentos, con lo cual se consagra la noción de unas bases neurales distinguibles para los talentos cognitivos (17,20,37).
La correspondencia entre las diferencias sexuales en la organización zonal encefálica y las habilidades cognitivas distintivas, en ambos géneros, emerge por tanto con asiduidad aunque de forma moderada. En un meticuloso estudio efectuado en un centenar de jóvenes universitarios madrileños (11) se constató que la ventaja volumétrica del cerebro de los varones, sobre el de las féminas, no muestra relación alguna con las medidas de agudeza cognitiva global. Aunque no hubo diferencias en el talento cognitivo de chicas y chicos, se detectaron distingos en aptitudes específicas: una notoria ventaja femenina en tareas exigentes de razonamiento verbal y una considerable ventaja varonil en la rotación espacial de figuras. A ese nivel, el de los talentos específicos sí se constataron vinculaciones moderadas entre los tamaños de los segmentos cerebrales (sustancia gris y blanca) y los rendimientos intelectuales, con distinciones entre sexos. La ventaja masculina en tareas espaciales parece sólida y quizás esté relacionada con una mayor circuitería para el procesamiento simultáneo en el mapeo de la configuración del espacio personal y del entorno, tal como se explora en las pruebas de rotación mental de figuras o en los tests de navegación espacial. En cambio, las contrastadas ventajas femeninas en las pruebas de fluencia e inducción verbal y en diversas medidas de memoria parecer ser que descansan en un procesamiento secuencial optimizado (2,5,36,64). El enraizamiento firme de esas distinciones en talento cognitivo en propiedades singulares de la organización cerebral requerirá, sin embargo, un nivel de ponderación morfométrica y de análisis funcionales, durante los escaneos, mucho más precisos que las estimaciones preliminares que se han venido obteniendo hasta ahora. Los estudios sobre diferencias sexuales en núcleos, áreas y haces, así como en sistemas corticales y subcorticales progresan a buen ritmo y a pesar de que la variabilidad sea un acompañante inevitable, están incrementando el poso de hallazgos sólidos sobre los dimorfismos sexuales y su relación con los talentos en el cerebro humano.
Resistencias ante los distingos sexuales.
Larry Cahill, en un ensayo (12) sobre los frentes de avance en las pesquisas sobre los dimorfismos encefálicos en varones y mujeres avanzó, hace una década, que se repiten en este ámbito una serie de errores y confusiones de manera recurrente, hasta el punto de distorsionar enormemente el panorama. Las tres primeros malentendidos están vinculados entre sí, a saber: 1. persiste el convencimiento erróneo de que las diferencias entre géneros son más bien pequeñas en términos del rendimiento conductual y cognitivo, así como en sus posibles sustratos neurales; 2. esas tenues distancias en promedio, entre ambos sexos, denotarían sobretodo la influencia de casos extremos en la distribución de las poblaciones estudiadas; y 3. las diferencias intra-sexuales suelen ser mucho mayores que las intersexuales, con lo cual los distingos debieran considerarse triviales. La cuarta fuente de error deriva de considerar que todas las diferencias sexuales, cuando son sistemáticas, obedecen a la acción de las hormonas sexuales, lo cual implicaría que los cerebros masculinos y femeninos son idénticos en esencia, salvo en el detalle de las fluctuaciones de las hormonas esteroideas. Un último frente de confusión deriva de la noción errónea de que cuando no hay manera de acotar una diferencia sexual, para un atributo o rendimiento cognitivo concreto, de ello cabe deducir que no habrá, por definición, diferencias ligadas al sexo en los sistemas neurales que lo regulan (12).
Aunque todas esas nociones han sido refutadas una y otra vez, perviven incluso entre los biólogos y los neurocientíficos que trabajan en ámbitos alejados de los distingos asignables al sexo. No remiten, probablemente, debido a la carga “polémica” que arrastra cualquier indagación enfocada a deslindar diferencias entre géneros. De todos modos, la necesidad de dilucidar los orígenes de la variabilidad debida al sexo no ha cesado de aumentar, aunque sólo sea por la urgencia de desbrozar los mecanismos hay detrás de las considerables disparidades en la incidencia de enfermedades neurológicas y psiquiátricas entre ambos sexos. Ese es un hiato muy considerable y conocido de antiguo, pero como algunas de esas anomalías son padecimientos no precisamente triviales del sistema nervioso y afectan a enormes sectores de población (afecciones dolorosas crónicas, trastornos depresivos y esquizofrénicos, conductas auto-lesivas, la demencia de Alzheimer y otros procesos neurodegenerativos, trastornos del sueño, los autismos y otras anomalías génico-madurativas en la infancia y adolescencia), la investigación neuroquímica y molecular orientada a deslindar inductores diferenciales en esos síndromes, así como factores potenciadores o cascadas protectoras, ligadas al sexo, conoció una expansión imparable (5,7,12). Expansión que ha crecido a gran ritmo para aprovechar, a fondo, las dianas diferenciales, en el cerebro de ambos sexos, que la genética molecular va abriendo sin cesar (32). Se están diseccionando, incluso, los “handicaps” asociados a portar modalidades de cerebro masculino “extremo” o cerebro femenino “extremo” (5,10).
Esa efervescencia en la investigación génica y neuroquímica sobre los distingos sexuales en el funcionamiento normativo y en la psicopatología permitirá, además, reconectar con ámbitos de la investigación psicológica donde la diferenciación sexual siempre estuvo presente. El campo donde eso fue un foco exploratorio habitual es el de la personalidad: tanto en las aproximaciones estadísticas para acotar rasgos “universales” del carácter (sociabilidad, neuroticismo, agresividad/placidez, laboriosidad, afición al riesgo y la novedad), como en el deslinde de atributos temperamentales más cercanos al umbral de la eclosión clínica (trastornos de la personalidad, trastornos afectivos), donde el distingo sexual siempre jugó un papel relevante (24,28,46). Pocas vacilaciones se han dado ahí: los datos sobre diferencias intersexuales en empatía, en reactividad afectiva, en ambición, en perfeccionismo, en espiritualidad, en observancia de normas, en dominancia o en permisividad y promiscuidad sexual, por poner algunos ejemplos, son abundantísimos. Aunque en alguno de esos rasgos predominen los aspectos afectivos sobre los cognitivos, en la mayoría de ellos se dan confluencias de ambos procesos que responden, a su vez, a las operaciones de una circuitería y unos engranajes mediadores que muestran sesgos intersexuales. Hay que dejar constancia, sin embargo, que algunas recopilaciones y análisis globales insistieron en que las diferencias en personalidad entre los dos sexos son más bien menores, sobre todo cuando se toman en consideración las grandes dimensiones del temperamento (28). Los estudios efectuados en muestras muy extensas y representativas de la población USA, extremando las cautelas metodológicas y midiendo un abanico amplio de rasgos de personalidad indican, no obstante, que las diferencias de temperamento entre hombres y mujeres son notables y que se acercan al hiato muy apreciable que se da en los perfiles combativos distintivos o en las conductas de protección y cuidado de la prole (19).
A partir de aquí voy a usar, tan sólo a modo de ejemplo, el rasgo de la agresividad diferencial entre ambos sexos porque es, quizás, el atributo donde se obtienen unas disparidades más apoteósicas: los hombres se llevan la palma en el uso de tácticas violentas pero la agresividad femenina no es nada banal, al tiempo que presenta cualidades y resortes peculiares.
La violencia (atenuada) femenina.
Las mujeres no son benignas. Aunque saben prodigar la ternura y abnegación que derivan de su andamiaje amamantador, cuidador y protector, acarrean unos resortes combativos nada despreciables. En los envites entre mujeres o en las cuitas con los hombres, la agresividad femenina despliega una considerable versatilidad y eficacia. Ocurre, sin embargo, que suele pasar más desapercibida por la aparatosidad y la asiduidad de los percances desencadenados por ellos (3,4,9,58,61). Los varones acaparan de una manera tan ostentosa la crónica de las brutalidades y atrocidades que a menudo parece como si la violencia femenina fuera tenue o nula; como si las féminas estuvieran infradotadas para el comportamiento dañino. Y no es así. Hay que constatar, eso sí, que las comparaciones sobre incidentes lesivos son taxativas: en el registro de homicidios, de asaltos con violencia física y de robos con intimidación suele haber nueve hombres implicados por cada mujer. Esa tremenda desproporción se repite en todas las sociedades tanto si han alcanzado estadios avanzados de desarrollo como si viven en hábitats primitivos. Y valen tanto para los incidentes graves como para los que se limitan a la disputa, el altercado o la trifulca sin mayores consecuencias. Hay, por consiguiente, una gran distancia en la proclividad dañina de ambos sexos aunque se descuenten las violaciones sexuales, un ámbito donde el monopolio masculino es casi incontestado (8,13,57).
La enormidad de esa diferencia oculta, sin embargo, la contribución femenina a la combatividad lesiva. No me estoy refiriendo, con ello, a los crímenes de gran resonancia protagonizados, de vez en cuando, por mujeres (terroristas o mafiosas destacadas, torturadoras profesionales, asesinas en serie, adolescentes que han martirizado a compañeras hasta la muerte o madres que han infringido padecimientos salvajes a sus hijos), porque esos episodios caen de lleno en aquel diez por ciento de alto voltaje criminal. De hecho, a pesar de que los registros sesgados con que solemos operar conduzcan a pensar que la proporción dañina diferencial, entre géneros, debe rondar el 99/1 o distancias todavía mayores, los datos reiteran el tozudo 9/1. Por otro lado, las oleadas de homicidios conyugales tienen ahora una repercusión tan estruendosa que en algunos momentos parece incluso como si el goteo de mujeres asesinadas por sus parejas llevara visos de alcanzar proporciones epidémicas. Aunque en esas bajas por violencia doméstica los porcentajes incriminatorios son también muy superiores para los hombres, la distancia entre ambos sexos es menor de lo que cabria suponer de hacer caso al voceo mediático. En cifras norteamericanas de las dos últimas décadas, el sesenta y dos por ciento de las muertes conyugales fueron debidas a los maridos y el treinta y ocho por ciento a las esposas. En España, los datos judiciales dan un setenta por ciento de maridos homicidas y un treinta por ciento de mujeres homicidas. Por tanto, la desproporción es muy alta pero no tan apabullante como suele pensarse (3,13,38,55).
Tipologias agresivas y habilidades de género. Cuando se registran con minuciosidad las agresiones de “baja intensidad” en la vida familiar, las mujeres se suelen llevar la palma en diversas medidas. En habilidades como los desplantes y los sarcasmos, las burlas y los gestos insidiosos, las conductas negativistas o la desatención vejadora hay predominancia de la agresividad femenina. Esos últimos datos concuerdan bien, por cierto, con otros muchos que han podido constatar una clara superioridad femenina en las aptitudes paradigmáticas de la inteligencia social. Esa ventaja en el talento empático la presentan las muchachas desde la infancia o la adolescencia más temprana y la saben administrar a lo largo de toda la vida adulta. Parece ser que las mujeres tienen un cerebro algo mejor equipado que el de los hombres para reconocer e interpretar los sentimientos de los demás y aprovechan esa superioridad para lastimar, cuando conviene, recurriendo a dardos verbales o gestuales que impactan, de lleno, en la línea de flotación de la autoestima del contrincante (33).
Ahora bien, en las cuitas entre hombres y mujeres debe contrastarse esa diferencia en talentos con la desproporción en poderío físico entre los dos sexos. Las mujeres tienen, por término medio, una corpulencia inferior en un 10-20% a la de los varones que puede, a su vez, resultar en una desventaja del 50% en fuerza muscular. Ese es un hándicap basal estentóreo que es probable que jugara un papel determinante en el reparto de roles y dominancias a lo largo del itinerario de nuestros ancestros. Ahora cuenta menos gracias al progreso moralizante y antiagresivo derivado, sobre todo, de los avances en tecnología (véanse las series de datos ilustrativos en 48). De todos modos, aquella discrepancia en poderío físico sigue siendo relevante porque en circunstancias de litigio grave confiere una ventaja radical. De ahí que los varones recurran con más frecuencia a la violencia física mientras que las mujeres usan otras tácticas lesivas. En las cuitas entre mujeres, sin embargo, aunque las sutilezas combativas aparecen con todo lujo de ardides no es excepcional que la escalada de paso al ataque físico (el “hooliganismo”, por ejemplo, es común entre jugadoras y aficionadas al fútbol femenino así como en otros esparcimientos gimnásticos).
Hay, pues, diferencias sustanciales entre la combatividad masculina y la femenina. Los hombres se apuntan a las tácticas que implican violencia física con mucha mayor asiduidad que las mujeres. El mecanismo que sustenta esa diferencia es la distancia en el armamento físico y los engranajes neuroendocrinos del empuje competitivo que tienen a su disposición ambos sexos. Por el contrario, en las tácticas que implican combatividad verbal, gestual o indirecta (la dirigida a los intereses, el estatus o la reputación de los rivales), los rendimientos andan muy igualados y en algunas habilidades combativas vinculadas a la cognición social las mujeres superan claramente a los hombres (3,4,6,13,55,61).
¿Cerebros combativos distintivos?. Esas diferencias en combatividad no pueden asignarse, por el momento, a una organización distintiva para los dos sexos en las estructuras y circuitos que conforman la trama íntima del cerebro combativo. Por lo que sabemos, las regiones neurales encargadas de dar curso a la orquestación fisiológica y comportamental de las salidas agresivas no presentan dimorfismos sexuales distinguibles (41,42). Hay que indicar, sin embargo, que se trata de engranajes primitivos del cerebro afectivo que son de difícil acceso y que no han sido abordados todavía con las técnicas de escaneo o de neuromorfología zonal más resolutivas, que ya han comenzado aplicarse en roedores y en simios. Las diferencias entre géneros son notorias, en cambio, en algunos sistemas subcorticales (amígdala, hipotálamo) que se encargan de orquestar reacciones de la combatividad emotiva como la ira o la hostilidad. Se ha llegado a vincular, incluso, la mayor presencia de trastornos de personalidad antisocial, en los varones, a la reducción de sustancia gris que presentan los individuos que acarrean esas psicopatías con tintes virulentos, en algunas regiones ventrales, mediales y orbitales de la corteza prefrontal. Existen datos incipientes que han corroborado ese nexo entre déficits zonales en sustancia gris prefrontal y la asocialidad diferencial, entre sexos, en muestras normativas y sin contacto alguno con el sistema judicial o el clínico (49).
Donde sí hay distingos sexuales firmes es en el armamentario neuroendocrino que actúa en esas regiones cerebrales para modular su trabajo. Me refiero, con ello, a las sustancias que procedentes de la periferia corporal o elaboradas en el propio cerebro trabajan en esas zonas neurales para facilitar o inhibir el empuje competitivo o los brotes agresivos. Las hormonas sexuales son los primeros protagonistas a tener en cuenta. Los andrógenos actúan como facilitadores de la combatividad y la dominancia, y las cifras circulantes son muy diferentes en hombres y en mujeres aunque presenten una enorme variabilidad dentro de cada sexo. Ya se tienen datos firmes que confirman que las mujeres dadas a la combatividad física, la dominancia y los esparcimientos que conllevan riesgo físico elevado presentan unas cifras androgénicas peculiares (18,39,43,44,47). Es decir, que las féminas de temple más ambicioso y temerario poseen un sesgo andrógino a pesar de la ausencia de glándulas testiculares. Por otra parte, las oscilaciones hormonales prototípicas del ciclo menstrual generan una reactividad cambiante en el estado de ánimo, que se acompaña de variaciones del umbral de irritabilidad ante los percances. Y la cosa no acaba ahí, por supuesto. Hay otras muchas sustancias que modulan las salidas agresivas, promoviendo o frenando la combatividad (45).
Las hormonas de alarma/estrés (las del eje corticoideo, la adrenalina, la noradrenalina, la vasopresina) juegan un papel muy relevante en los envites competitivos y hay diferencias sexuales consistentes. Así, por ejemplo, existen datos que señalan que las adolescentes con una conducta reiteradamente disruptiva no sólo comparten con los chicos más rebeldes un incremento en los perfiles testosterónicos sino unas cifras consistentemente bajas de cortisol (la hormona diana del estrés dañino). Por otra parte, las diferencias sexuales en el funcionamiento serotonérgico u opioideo central están siendo mapeadas con precisión y ejercen influencia en los estilos competitivos de ambos sexos porque hay multitud de hallazgos que las relacionan con la modulación inhibitoria de la agresividad. Y todavía hay más. Los niveles de oxitocina modulan la proximidad, el gozo y la dependencia afectiva y eso implica, de ordinario, una restricción para las expresiones agresivas. Comienza a haber datos, asimismo, de que la vasopresina central es un inductor agresivo relacionado con la posesividad celosa en animales que establecen fusiones de pareja, así como en humanos. Los celos constituyen un dinamizador emotivo con trazos distintivos en función del género y pródigo para los conflictos sentimentales graves. El empeño en dibujar unos perfiles neuroendocrinos que sustenten los rasgos de agresividad distintiva, en hombres y mujeres va mucho más allá, por descontado, de lo que sugiere este resumen apresurado (42,45).
Combatividad y entorno cultural. ¿Todo depende, al fin y al cabo, de las prescripciones y las modulaciones de orden biológico? ¿Debemos asignar las diferencias en el talante combativo entre hombres y mujeres a una descripción pormenorizada de las cascadas neuroendocrinas y la circuitería neural de la agresividad ? ¿Dónde quedan las influencias de la cultura patriarcal, de los estilos educativos distintivos, de las desigualdades y los “techos de cristal” discriminatorios en las sociedades “igualitarias”?. Pues sí, ese es el mensaje: las diferencias sexuales en combatividad deben enraizarse, en primerísimo lugar, en los engranajes neuroquímicos porque no hay frontera más nítida que el sexo en la diferenciación biológica. No sólo hay un cromosoma entero a su servicio sino un vasto cortejo de señales moleculares dedicadas a la cristalización de unas morfologías externas y unos dispositivos internos meridianamente distintos. Eso incluye el cerebro, lugar donde se cuece en última instancia la expresión de un rasgo tan conspicuo del carácter como la agresividad (62).
He repasado datos suficientes para corroborar que las mujeres no son benignas. No lo son aunque esté archidemostrado que sus estilos combativos ocasionan muchísimas menos bajas que los de sus congéneres masculinos. Las influencias culturales tienen, no obstante, una potencia insoslayable porque pueden alterar esa morbilidad diferencial. Usaré dos ejemplos. Ultimamente se viene registrando una ligera pero firme tendencia al alza de la combatividad femenina, en todos los frentes, que está erosionando aquella ominosa desproporción de 9/1 del daño varonil respecto del femenino. Esos hallazgos son tentativos pero tanto en las agresiones que conducen al encarcelamiento como en los alborotos y encontronazos de menor importancia aquella desproporción está mermando hasta acercarse al 8/2. Esa creciente presencia femenina en la combatividad lesiva habrá que asignarla, en principio, a variaciones culturales recientes que permiten que emerja una malignidad frenada por las constricciones sociales (38). Hay otros datos concordantes: en las adolescentes muy disruptivas (las que prodigan los episodios de novillos, escapadas del hogar, hurtos, gamberrismo, peleas con los compañeros, crueldad para con los animales o las personas débiles), las distancias entre chicos y chicas son inferiores a lo esperado. La incidencia promedio de esa anomalía comportamental es del 8% para las muchachas y del 12% para los chicos. Ese peculiar hiato combativo es, por tanto, mucho más modesto. Una buena parte de esas adolescentes culminan una socialización satisfactoria en los años subsiguientes, pero una porción no desdeñable persiste con la conducta conflictiva que extienden hasta la juventud y las etapas adultas.
El siguiente ejemplo concierne a la influencia de las horas de televisión durante la infancia, adolescencia y primera juventud sobre la conducta agresiva posterior. La evidencia más sólida procede de un trabajo longitudinal efectuado en una muestra representativa de familias del estado de Nueva York (30). A lo largo de 25 años evaluaron a los hijos/hijas de 707 familias mayoritariamente blancas y católicas, con una edad de cinco años al inicio del seguimiento. Se obtuvieron múltiples datos en cuatro ocasiones distintas a lo largo del periodo y la evaluación final, en el 2000, incluyó no sólo las respuestas de hijos y madres a los cuestionarios sino datos de los archivos policiales del FBI. Los resultados indicaron que el alto consumo de televisión durante la adolescencia (a los 14 años) se vinculaba con una mayor incidencia de conductas agresivas a los 16 y los 22 años, con cifras superiores para los muchachos. Esa vinculación, se reprodujo al relacionar las horas dedicadas a ver TV a los 22 años con la conducta agresiva medida a los 30 años, pero en éste último caso el efecto era más potente para las chicas, hasta el punto de igualar los resultados de los varones. En conjunto, esos hallazgos revelaron que existe un subgrupo de jóvenes de ambos sexos en los que coincide un alto consumo televisivo con todo tipo de comportamientos disruptivos y delictivos, sin que pueda discernirse una direccionalidad clara en un cóctel que se retroalimenta (temple agresivo + alto consumo televisivo). Hay que decir que ese efecto se mantenía, asimismo, cuando se aislaba la influencia del nivel socioeconómico deprimido, el abandono infantil, los abusos precoces, el consumo de sustancias ilegales y otras variables contextuales que predicen agresividad.
Conclusión: poder femenino y masculino.
Hay que destacar que en las investigaciones sobre dimorfismos sexuales en humanos se ha podido avanzar ya desde el estadio descriptivo y correlacional, hasta los experimentos dirigidos. Así, administrando dosis bajas de testosterona las mujeres pierden capacidad para el reconocimiento emotivo y para las aptitudes relacionadas con la empatía cognitiva. Es decir, disminuye su ventaja en atributos nucleares de la inteligencia social donde, de ordinario, suelen superan con creces a los varones. Lo mismo ocurre con la hostilidad, la aversión al riesgo y el temor ante las amenazas físicas, en consonancia con cambios funcionales en el trabajo de la circuitería cerebral moduladora de la agresividad (63). Y ha podido demostrarse también que la cautela femenina ante las decisiones financieras arriesgadas puede modificarse mediante una potenciación androgénica (50). En conjunto, por consiguiente, se han obtenido pruebas de que un sesgo neuroendocrino inducido, de manera transitoria y actuando sobre engranajes focales y distintivos, es capaz de cambiar el estilo cognitivo y el afectivo característicos de uno de los géneros, para acercarlo al del sexo opuesto.
Todo ello robustece la aproximación que he seguido aquí y sirve para ilustrar, con solvencia, una aserción que buena parte de los datos apoyan: hombres y mujeres disponen de circuitos neurales y arsenales neuroendocrinos para cooperar al tiempo que competir, con denuedo, en escenarios sociales muy complejos y siempre cambiantes. Los más complejos, con toda seguridad, de todo el reino animal. Los rendimientos, por ambos lados, son espléndidos y parecidos, en general, con algunos perfiles y especializaciones optimizadas. En definitiva, que los hombres y las mujeres usan unos cerebros y unos cócteles neuroquímicos parcialmente distintos para intentar alcanzar cotas exigentes y, en la medida de lo posible, similares, en todos los ámbitos de las cuitas y desvelos humanos, incluyendo las batallas por el poder social (60). La tarea de los neurocientíficos consiste en ir desbrozando las rutas comunes y las distintivas, en ambos géneros, sin dejarse contaminar en demasía por el ruido de los envites incesantes por conseguir preeminencias o paridades políticas o sociales (54,56).
[1] Las polémicas sobre sutiles factores culturales que podrían conducir a techos diferenciales en algunos ámbitos disciplinares no cesan (27) y han dado lugar a intercambios sugerentes. Véase, por ejemplo, los datos confrontados de (16) y (22), sobre el efecto de las expectativas en cuanto al talento natural diferencial.
Referencias.
- Allen JS, Damasio H., Grabowski TJ, Bruss J. and Zhang W (2003) Sexual dimorphism and asymmetries in the gray–white composition of the human cerebrum, Neuroimage 18, 880–894 (2003).
- Andreano JM and Cahill L (2009) Sex influences on the neurobiology of learning and memory, Learning andMemory, 16, 248-266.
- Archer, J. (2004). Sex differences in aggression in real-world settings: a meta-analytic review. Review of General Psychology, 8, 291–322.
- Archer J (2009) Does sexual selection explain human sex differences in agression?, Behavioral and Brain Sciences, 32, 249-311.
- Baron-Cohen S, Knickmeyer RC and Belmonte MK (2005) Sex differences in the brain: implications for explaining autism, Science, 310, 819-823.
- Benenson JF, Markovits H, Thompson ME and Wrangham RW (2011) Under threat of social exclusion, females exclude more than males, Psychological Science, 22, 4, 538-544
- Berkley KJ (2003) Sexual differences in pain, Behavioral and Brain Sciences, 20, 371-380.
- Bettencourt BA and Miller N (1996) Gender differences in aggression as a function of provocation: a meta-analysis, Psychological Bulletin, 119, 422-447.
- Bjorquist K, Osterman K, Kaukiainen A and Logerspetz KMJ (1999) Concomitants of physical, verbal and indirect aggression, Aggressive Behavior, 25, 1, 35.
- Bremser JA and Gallup GG (2012) From one extreme to the other: negative evaluation anxiety and disordered eating as candidates for the extreme female brain, Evolutionary Psychology, 10, 457-486.
- Burgaleta M, Head K, Alvarez-Linera J, Martínez K,Escorial S, Haier R and Colom R (2012) Sex differences in brain volume are related to specific skills, not to general intelligence, Intelligence, 40, 60-68.
- Cahill L (2006) Why sex matters for neuroscience, Nature Reviews Neuroscience, 7, 6, 477-484.
- Campbell A (1999) Staying alive: evolution, culture and women’s intra-sexual aggression, Behavioral and Brain Sciences, 22, 203-252.
- Ceci SJ and Williams WM (2010a) The mathematics of sex: how biology and society conspire to limit talented women and girls, New York: Oxford University Press.
- Ceci SJ. And Williams WM (2010b) Sex differences in math-intensive fields, Current Directions in Psychological Science, 19, 275–279.
- Ciampiani A and Leslie SJ (2015) Response to comment on expectations of brilliance underlie gender distributions across academic disciplines, Science, 349, 6246, 391-c.
- Clayden JD, Jentschke S, Muñoz M, Cooper JM, Chadwick MJ, Banks , Clark ChA and
Vargha-Khadem F (2012) Normative Development of White Matter Tracts: Similarities and Differences in Relation to Age, Gender, and Intelligence, Cerebral Cortex, 22, 8, 1738-1747.
- Dabbs JM, Barry Ruback R, Frady RL, Hopper ChH and Sgoutas DS (1988) Saliva testosterone and criminal violence among women, Personality and Individual Differences, 9, 2, 269-275.
- Del Giudice M, Booth T and Irving P (2012) The distance between Mars and Venus: measuring global sex differences in personality, PLOsOne, 7, 1, e29265.
- Escorial S, Román FJ, Martínez K, Burgaleta M, Karama Sh and Colom R (2015) Sex differences in neocortical structure and cognitive performance: a surface-based morphometry study, Neuroimage, 104, 355-365.
- Georgiadis JR, Kringelbach LM and Pfaus JG (2012) Sex for fun: a synthesis of human and animal neurobiology, Nature Reviews Urology, 9, 486-498.
- Ginter DK and Khan Sh (2015) Comment on Expectations of brilliance underlie gender distributions across academic disciplines, Science, 349, 6246, 391-b.
- Goldstein JM, Seidman LJ, Horton NJ, Makris N, Kennedy DN, Caviness VS, Faraone SV and Tsuang MT (2001) Normal sexual dimorphism of the adult human brain as assessed by in vivo magnetic resonance imaging, Cerebral Cortex, 11, 490-497.
- Gomà M (2001) Prosocial and antisocial aspects of personality in women: a replication study, Personality and Individual differences, 30, 1401-1411.
- Gur RC, Turetsky BI, Matsui M, Yan M, Bilker W, Hughett P and Gur RE (1999) Sex differences in brain gray and white matter in healthy young adults: correlations with cognitive performance, The Journal of Neuroscience, 19, 10, 4065-4072.
- Haier RJ, Jung RE, Yeo RA, Head K and Alkired MT (2005) The neuroanatomy of general intelligence: sex matters, Neuroimage, 25, 320-327.
- Hoffman M, Gneezy U and List JA (2011) Nurture affects gender differences in spatial abbilities, PNAS, 108, 36, 14786-14788.
- Hyde JS (2005) The gender similarities hypothesis, American Psychologist, 60, 581–592.
- Hyde JS and Mertz JE (2009) Gender, culture and mathematics performance, Proceedings of the National Academy of Sciences USA, 106, 8801–8807
- Johnson JG, Cohen P, Smailes EM, Kasen, S and Brook J (2002) Television viewing and aggressive behavior duing adolescence and adulthood, Science, 295, 2468-2471.
- Jones CM, Braithwaite VA, Healy SD (2003) The evolution of sex differences in spatial ability, Behavioral Neuroscience, 117, 403-411.
- Jung Kang H et al (2011) Spatio-temporal transcriptome of the human brain, Nature, 478, 483-489.
- Kaukiainen A, Bjorkquist K, Osterman K and Lagerspetyz KMJ (1996) Social intelligence and empathy as antecedents of different types of aggression, in CG Ferris, and Th Grisso (Eds) Understanding aggressive behavior in children, Annals New York Academy of Sciences, Vol. 794, 364-66.
- Kimura D (2000) Sex and cognition Cambridge, MA: MIT Press.
- Lenroot RK, Gogtay N, Greenstein DK,Wells EM, Wallace GL,Clasen LS, Blumenthal JD, Lerch J, Zijdenbos AP, Evans AC, Thompson PM and Giedd JN (2007) Sexual dimorphism of brain developmental trajectories during childhood and adolescence, Neuroimage, 36, 1065-1073.
- Lohman D and Lakin J (2009) Consistencies in sex differences on the cognitive abilities test across countries, grades, test forms and cohorts, British Journal of Educational Psychology, 79, 389–407
37. Luders E, Narr KL, Thompson PM, Rex DE, Jancke L, Steinmetz H and Toga AW (2004) Gender differences in cortical complexity, Nature Neuroscience, 7, 799 – 800
- Magdol L, Moffit TE, Caspi A, Newman DL, Fagan J and Silva PA (1997) Gender differences in partner violence in a birth cohort of 21-year-olds: bridging the gap between clinical and epidemiological approaches, Journal of Counsulting and Clinical Psychology, 65, 68-78.
- Maras A, Laucht M, Gerdes D, Wilhelm C, Lewicka S, Haack D, Malisova L and Schmidt MH (2003) Association of testosterone and dihydrotestosterone with externalizing behavior in adolescent boys and girls, Psychoneuroendocrinology, 28, 932–940.
- Nelson D and Brammer C (2010) A national analysis of minorities in science and engineering faculties at research universities (http://chem.ou.edu/*djn/diversity/faculty)
- Nelson RJ and Trainor BC (2007) Neural mechanisms of aggression, Nature Reviews Neuroscience, 8, 536-546.
- Niehoff D (1999) The biology of violence: how understanding the brain, behavior and environment can break the vicious cycle of aggression, New York: Free Press.
- Pajer KA (1998) What happens to “bad” girls?: a review of the adult outcomes of antisocial adolescent girls, American Journal of Psychiatry, 155, 7, 862-870.
- Pajer K, Gardner W, Rubin RT, Perel J and Neal S (2001) Decreased cortisol levels in adolescent girls with conduct disorders, Archives of General Psychiatry, 58, 3, 297-302.
- Pfaff DA (2011) Man and woman: an inside sotory, NY: Oxford University Press.
- Pintzinger NM, Pfabigan DM, Pfau L, Kryspin-Exner I and Lamm C (2017) Temperament differentially influences early information processing in men and women: preliminary electrophysiological evidence of attentional biases in healthy individuals, Biological Psychology, 122, 69-79.
- Pope HJ, Kouri EM and Hudson JI (2000) Effects of supraphysiologic doses of testosterone on mood and aggression in normal men: a randomized controlled trial, Archives of General Psychiatry, 57, 2, 133-140.
- Pinker S (2011) The better angels of our nature: why violence has declined, N.Y.: Viking Penguin.
- Raine A, Young Y, Narr KL and Toga W (2011) Sex differences in orbitofrontal gray as a partial explanation for sex differences in antisocial personality, Molecular Psychiatry, 16, 227-236.
- Sapienza P, Zingales L and Maestripieri D (2009) Gender differences in financial risk aversion and career choices are affected by testosterone, PNAS, 106, 15268–15273.
- Shah NM, Pisapia DJ, Maniatis, S Mendelsohn MM, Nemes A and Axel R (2004) Visualizing sexual dimorphism in the brain, Neuron, 43, 313–319
- Sommer IE, Aleman A, Bouma A, Kahn RS (2004) Do women really have more bilateral language representation than men? A meta-analysis of functional imaging studies, Brain, 127, 1845–1852.
- Sowell ER, Peterson BS, Kan E, Woods RP, Yoshii J, Bansal R, Xu D, Zhu H, Thompson PM and Toga AW (2007) Sex differences in cortical thickness mapped in 176 healthy individuals between 7 and 87 years of age, Cerebral Cortex, 17, 1550-1560.
- Stern Ch (2016) Undoing insularity: a small study of gender sociology’s big problem, Econ Journal Watch, 13, 3, 452-466.
- Stockley P and Campbell A (2013) Female competition and aggression: interdisciplinary perspectives, Philosophical Transections of the Royal Society-B, 368, 20130073.
- Stoet G and GearyDC (2015) Sex differences in academic achievement are not related to political, economical or social equality, Intelligence, 48, 137-151.
- Thornhill R and Palmer CT (2000) Why men rape, The Sciences, 1, 30-36.
- Tobeña A (2001) Anatomia de la agresividad humana, Barcelona: Galaxia Gutenberg.
- Tobeña A (2006) El cerebro erótico, Madrid: La Esfera de los Libros.
- Tobeña A (2008) Cerebro y poder, Madrid. La Esfera de los Libros.
- Tobeña A (2016) Neurologia de la maldad, Barcelona: Plataforma.
- Valla JM and Ceci SJ (2011) Can sex differences in science be tied to the long reach of prenatal hormones? Brain organization theory, digit ratio (2D/4D) and sex differences in preferences and cognition, Perspectives on Psychological Science, 6, 2, 134-146.
- Van Honk J, Schutter DJ, Bosa PA, Kruijtc AW, Lentjesd EG and Baron-Cohen S (2011) Testosterone administration impairs cognitive empathy in women depending on second-to-fourth digit ratio, PNAS, 108, 8, 3448-3452.
- Wittelson S, Beresh H and Kigar DL (2006) Intelligence and brain size in 100 postmortem brains: sex, lateralization and age factors, Brain, 129, 386-398.
[1] Las polémicas sobre sutiles factores culturales que podrían conducir a techos diferenciales en algunos ámbitos disciplinares no cesan (27) y han dado lugar a intercambios sugerentes. Véase, por ejemplo, los datos confrontados de (16) y (22), sobre el efecto de las expectativas en cuanto al talento natural diferencial.

Adolf Tobeña
Monográfico Mujeres fuertes, hombres frágiles
Departamento de Psiquiatria y Medicina Legal. Instituto de Neurociencias. Universidad Autónoma de Barcelona
Marzo de 2017